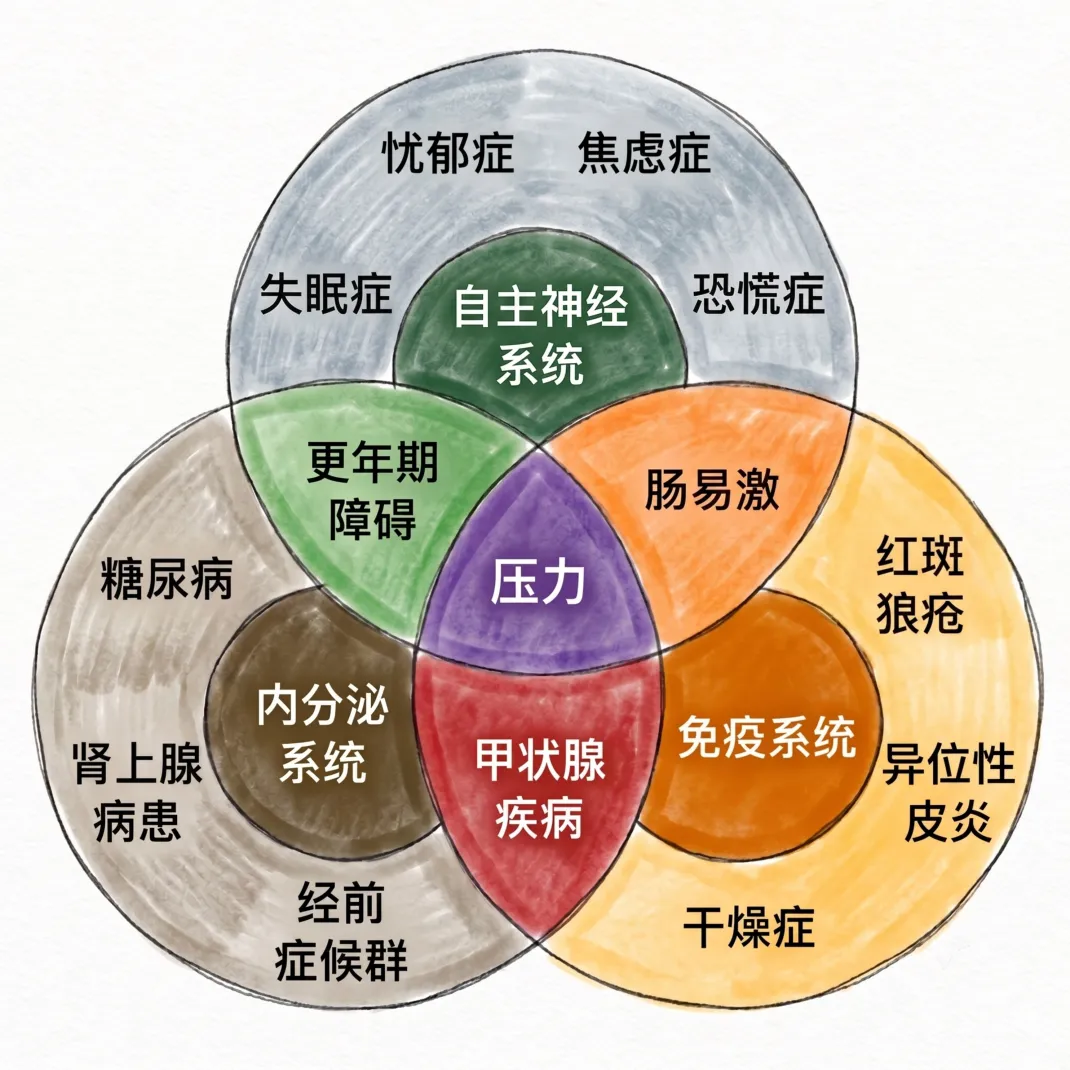
自主神经系统作为人体最重要的自动调控系统(身体的“自动驾驶系统”),其功能涵盖了从分子递质到系统行为的多个层次。交感神经和副交感神经在解剖上的差异——中枢起源不同、节前节后的长度比例各异、突触联系的弥散与局限之别——决定了它们在功能上“全局动员”与“局部微调”的互补优势。
两者在生理调控上有拮抗以保持平衡,也有协同以应对复杂情境;在病理状态下,无论是糖尿病长期侵蚀所导致的弥漫性自主神经损害,还是颈交感通路某一点的阻断所致的霍纳综合征,均彰显了自主神经精准调控的脆弱性。
从时间生物医学的角度审视,自主神经系统两大分支的功能转化,是人类在24小时昼夜周期中生存和适应的生物学基石。当昼夜调度受到干扰时,从心率失常到消化障碍,从血压失稳到情绪紊乱,全身多系统会随之失衡——这不仅是自主神经系统功能的一处“故障”,更是时间生物医学层面机体整体失调的信号。
以西医学为师,深入了解自主神经系统的解剖、组织学、作用机制、作用特点等。
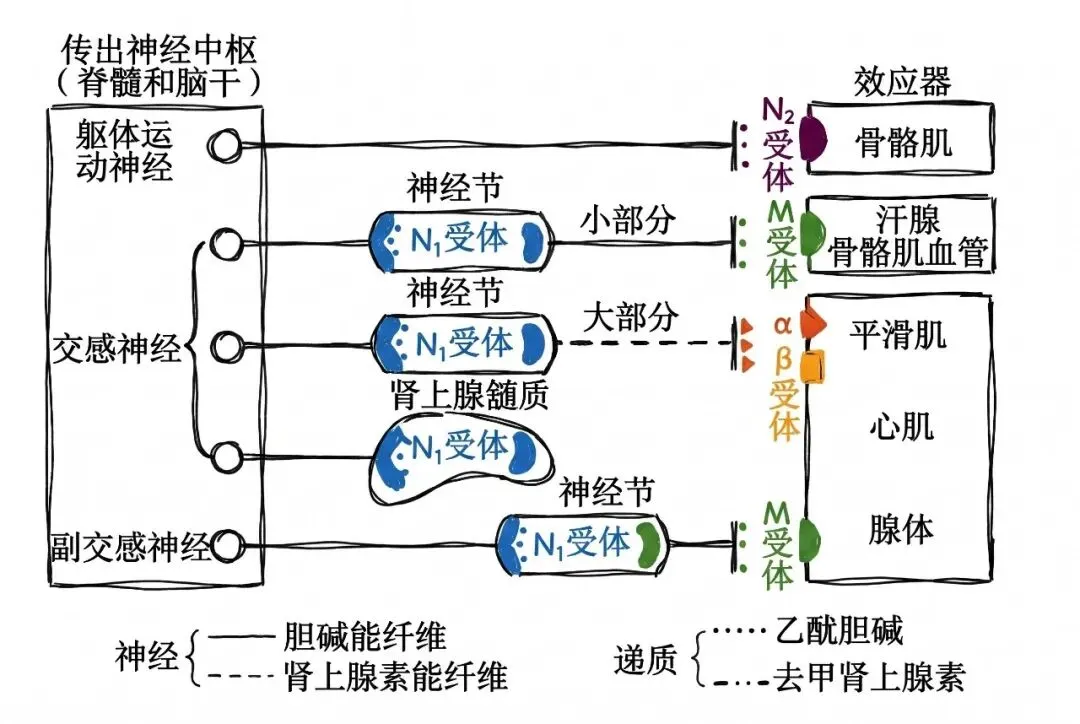
(1)中枢起源不同
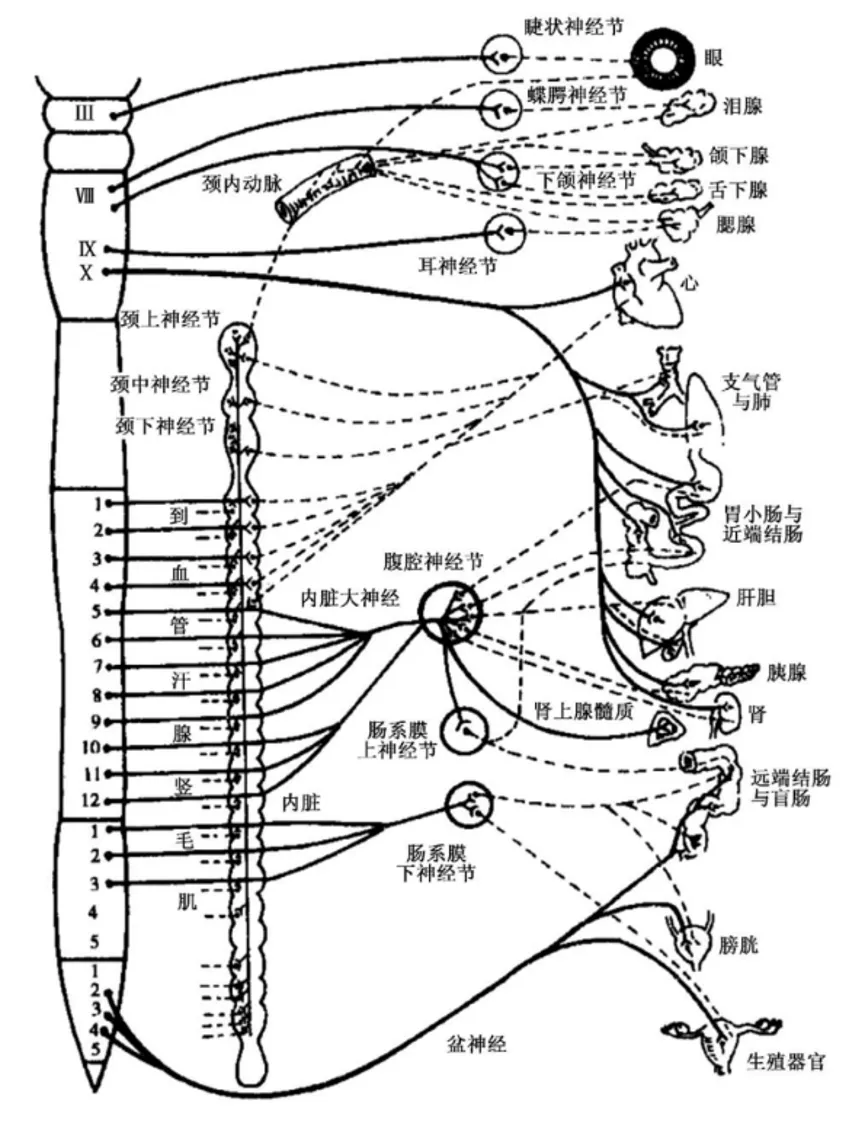
交感神经起源于脊髓胸腰段(T1~L3)的灰质侧角;副交感神经起源于脑干内副交感核和脊髓骶段(S2~S4)灰质(相当于侧角的部位)。
(2)节前纤维和节后纤维
自主神经由节前纤维和节后纤维组成,从中枢发出的自主神经纤维先进入外周神经节内换元,由节内神经元再发出纤维支配效应器(“换元”是自主神经和躯体神经的主要神经元结构差异)。交感神经节离效应器较远(前短后长),故节前纤维短而节后纤维长;副交感神经节离效应器较近(前长后短),有的神经节就在效应器官壁内,故节前纤维长而节后纤维短。
(3)双重神经支配
人体多数器官都接受交感神经和副交感神经双重支配(同时兴奋时,则占优势张力的神经的效应通常会显现出来),但交感神经的分布更广泛(皮肤和肌肉内的血管、一般的汗腺、竖毛肌、肾、肾上腺髓质仅有交感神经支配),几乎所有内脏器官都受它支配;而副交感神经的分布较局限,某些器官不受副交感神经支配。
(1)递质
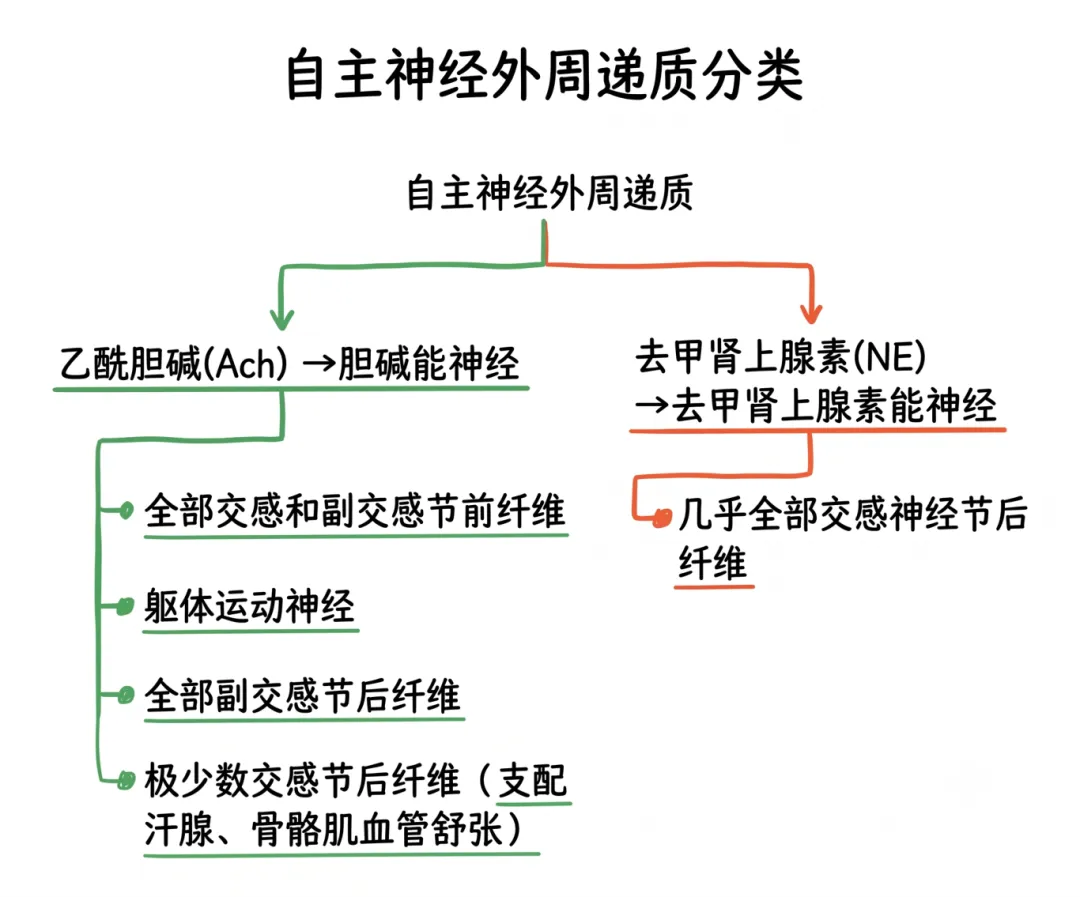
自主神经释放的递质属于外周递质,主要为乙酰胆碱(Ach)和去甲肾上腺素(NE)。凡末梢能释放乙酰胆碱的神经纤维称为胆碱能神经,胆碱能神经主要包括:①全部交感神经和副交感神经的节前纤维;②躯体运动神经、全部副交感神经的节后纤维;③极少数交感神经节后纤维,如支配汗腺分泌和骨骼肌血管舒张的神经纤维。凡末梢能释放去甲肾上腺素的神经纤维称为去甲肾上腺素能神经。去甲肾上腺素能神经则包括几乎全部交感神经节后纤维。
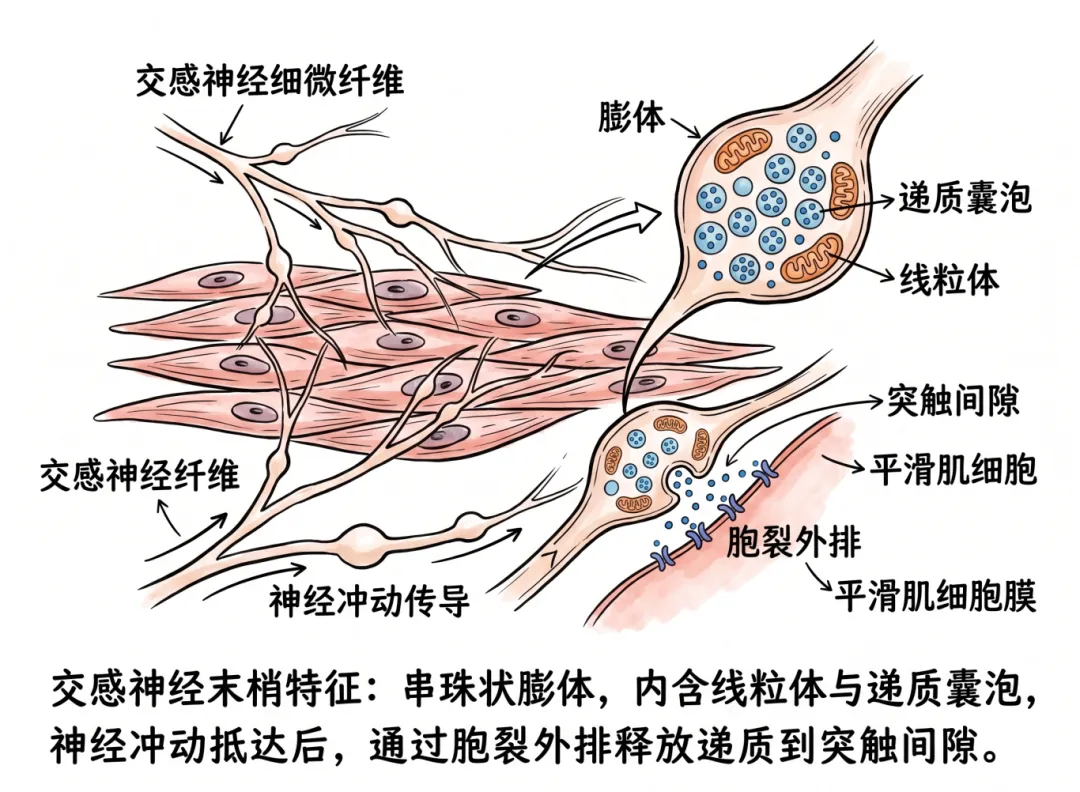
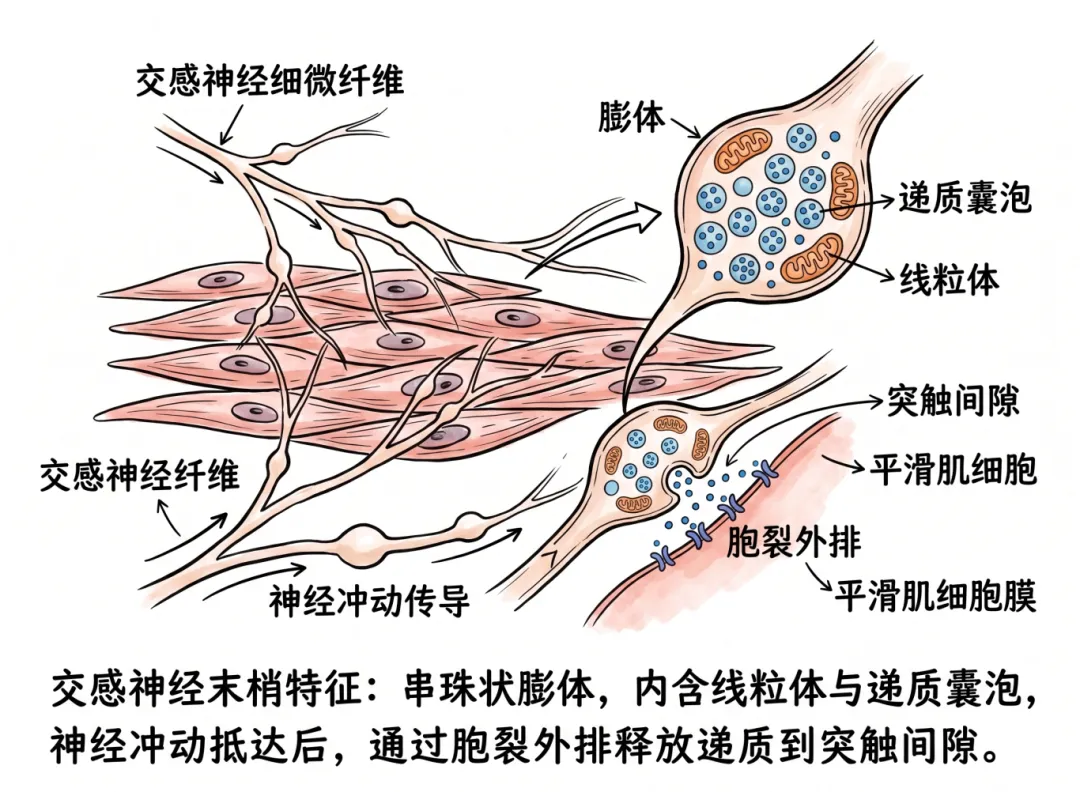
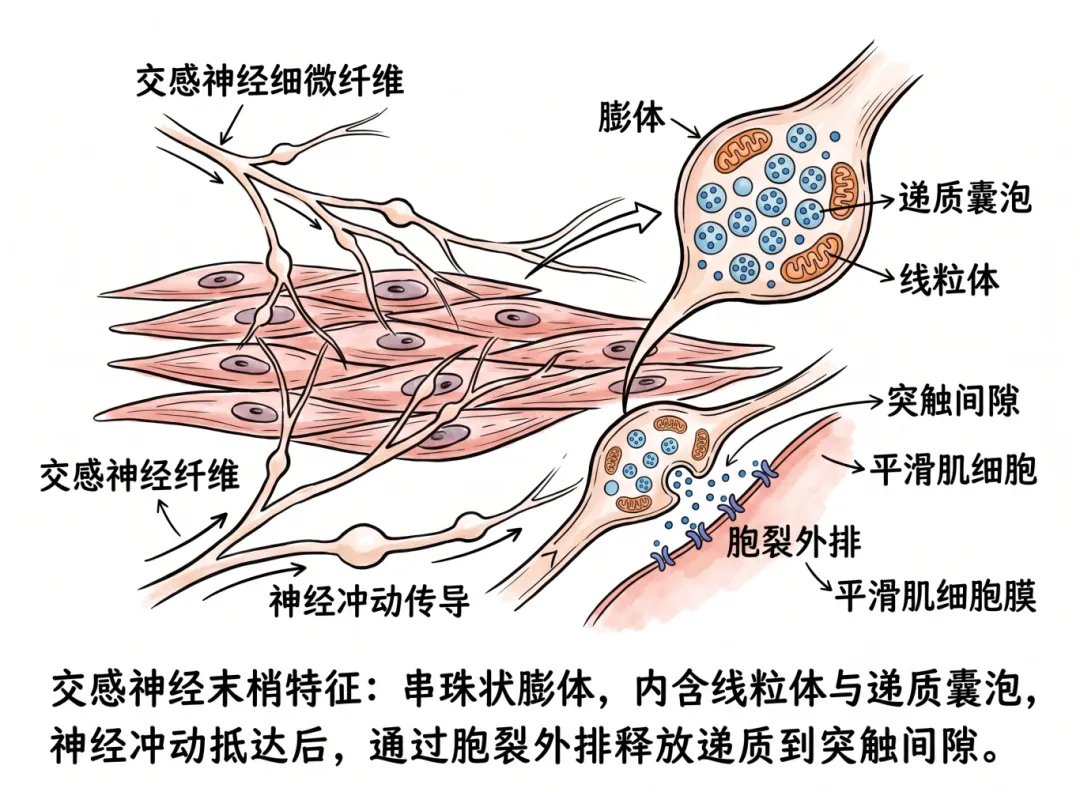
在组织学上,交感神经末梢显示出独特的结构特征——许多细微的神经纤维在平滑肌细胞之间分布,这些纤维上布有串珠状的膨胀部分,称为膨体,内含线粒体和储存递质的囊泡。当神经冲动抵达末梢时,以胞裂外排方式将递质释放入突触间隙。
(2)受体
自主神经节细胞和效应器细胞膜上都存在与递质相应的受体,可分为胆碱能受体和肾上腺素能受体。
【胆碱能受体】
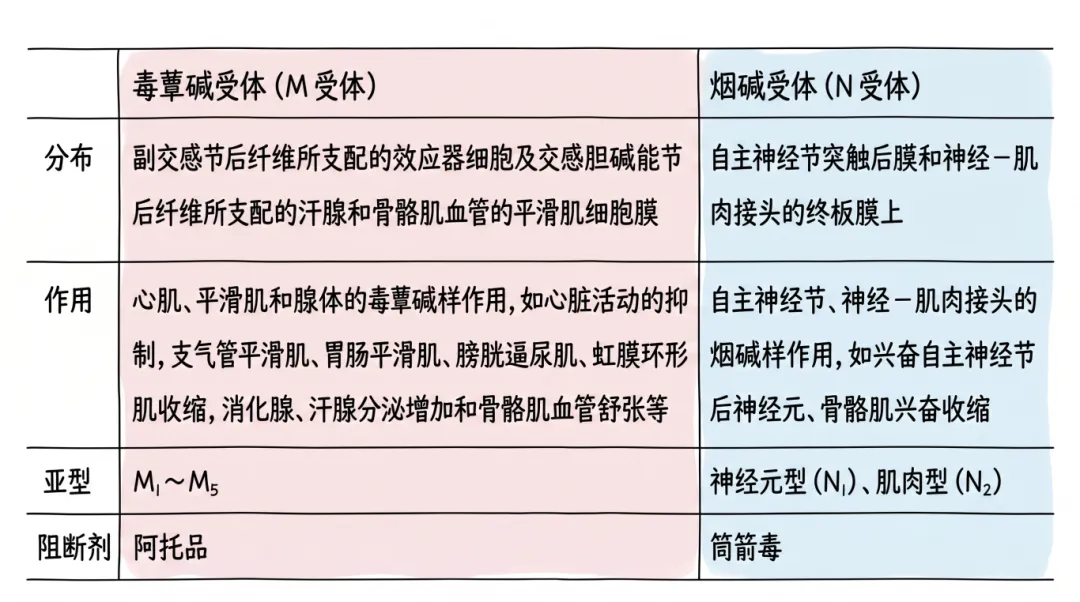
能与乙酰胆碱特异性结合的受体称为胆碱能受体,分为毒蕈碱受体和烟碱受体两类。它们存在于中枢神经系统内,因分别能与天然植物中的毒蕈碱和烟碱这两种生物碱相结合并产生不同生物效应而得名。
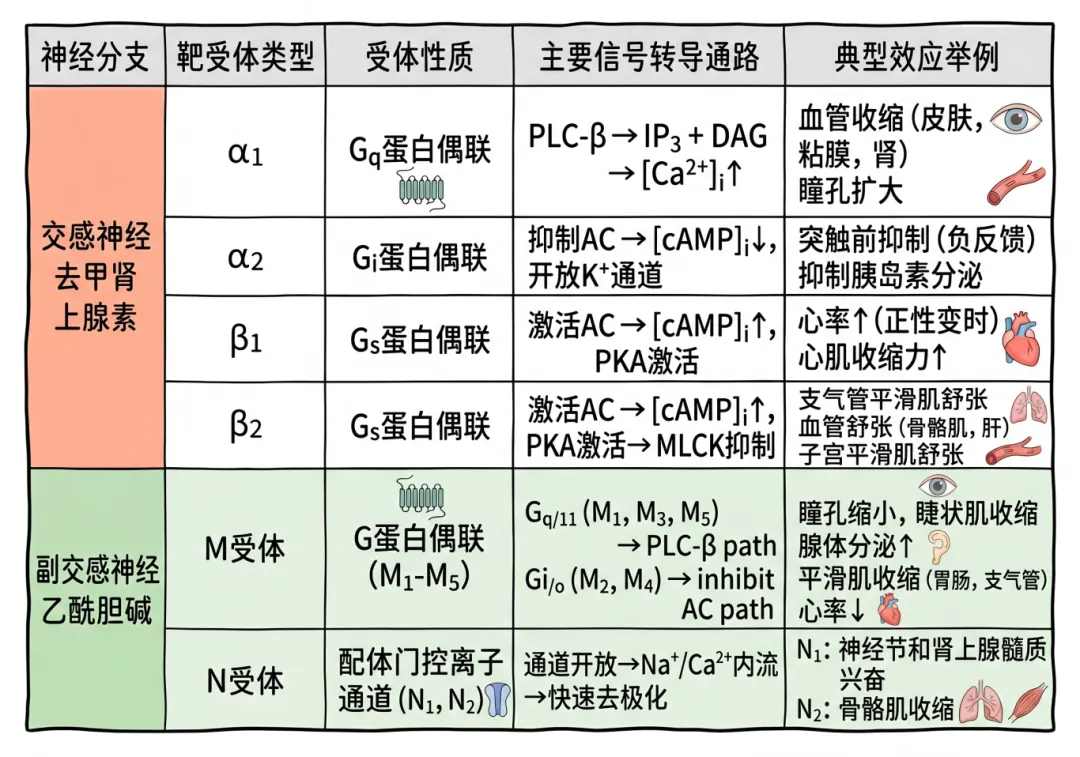
毒蕈碱受体(M受体):分布于几乎所有的副交感节后纤维所支配的效应器细胞及部分交感胆碱能节后纤维所支配的效应器(汗腺、骨骼肌血管的平滑肌细胞膜)上。已发现的M受体有五种亚型。乙酰胆碱与M受体结合后产生一系列自主神经效应,包括心脏活动的抑制,支气管平滑肌、胃肠平滑肌、膀胱逼尿肌、虹膜环形肌收缩,消化腺、汗腺分泌增加和骨骼肌血管舒张等。
烟碱受体(N受体):N受体分为两种亚型——N1受体(又称神经元型烟碱受体):位于自主神经节突触后膜上和肾上腺髓质嗜铬细胞上的受体;N2受体(又称肌肉型烟碱受体):存在于骨骼肌运动终板膜上的受体。两种受体都属于化学门控通道。小剂量乙酰胆碱能兴奋自主神经节后神经元,也可引起骨骼肌收缩,而大剂量乙酰胆碱则可阻断自主神经节的突触传递。
【肾上腺素能受体】
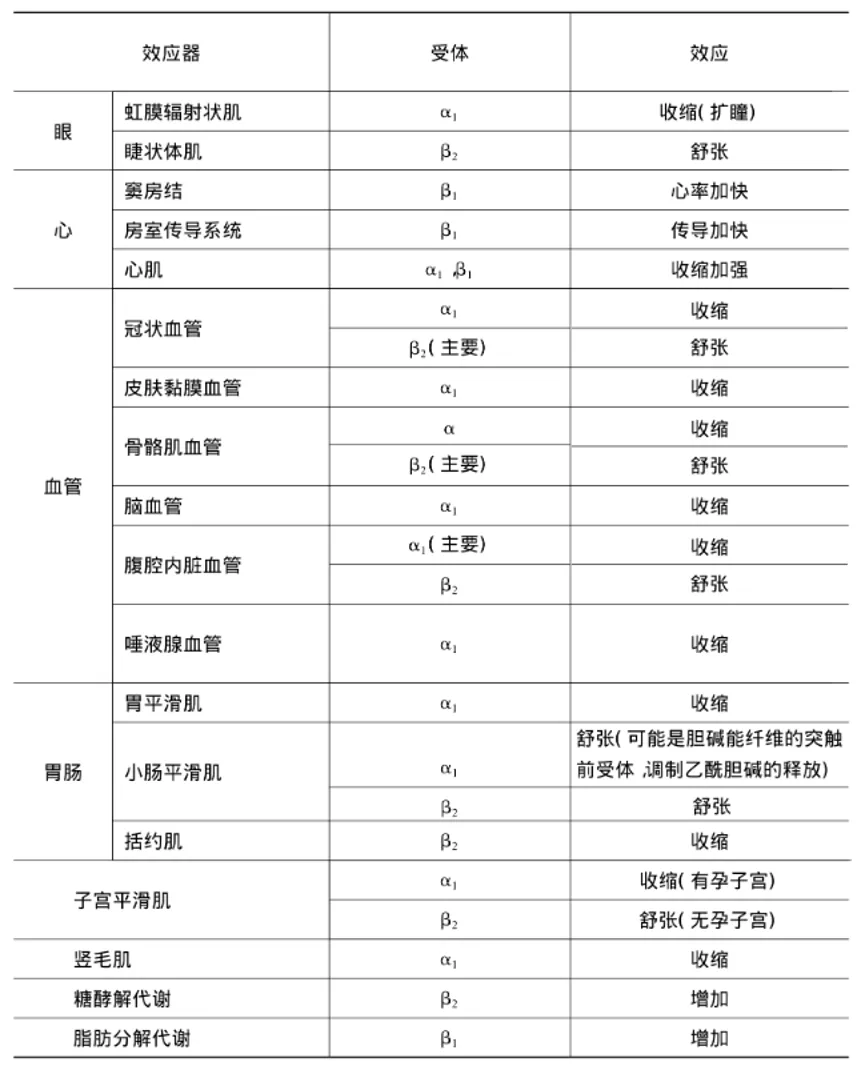
能与去甲肾上腺素结合的受体称为肾上腺素能受体,分为两类亚型和五种受体(α受体和β受体两类,α受体又可分为α1、α2,β受体也可再分为β1、β2、β3)。多数交感神经节后纤维支配的效应器细胞膜上都有肾上腺素能受体。但在具体的某一效应器官上,有的仅有α受体,有的仅有β受体,也有的兼有两种受体。在皮肤、肾、胃肠的血管平滑肌上以α受体为主,而骨骼肌和肝脏的血管则以β受体为主。α和β受体不仅与交感神经神经节后纤维释放的递质起反应,也对血液中的儿茶酚胺起反应。去甲肾上腺素对α受体的作用较强,对β受体的作用较弱;肾上腺素对α和β受体的作用都很强;异丙肾上腺素主要对β受体有强烈作用。
去甲肾上腺素与α受体(主要是α1受体)结合后产生的平滑肌效应主要是兴奋的,包括血管、子宫、虹膜辐射状肌等的收缩,但也有抑制性的,如小肠舒张。酚妥拉明为α受体阻断剂可消除去甲肾上腺素引起血管收缩、血压升高的作用。
去甲肾上腺素与β受体(主要是β2受体)结合后产生的平滑肌效应主要是抑制的,包括血管、小肠、支气管等的舒张,但与心肌β1受体结合产生的效应却是兴奋的。β3受体主要分布于脂肪组织,与脂肪分解有关。普萘洛尔(心得安)是β受体阻断剂(包括β1和β2受体)。阿替洛尔能阻断β1受体,而丁氧胺(心得乐)则主要阻断β2受体。
(3)传出神经功能
去甲肾上腺素能神经兴奋时,神经末梢释放去甲肾上腺素,引起心脏兴奋,皮肤、粘膜和腹腔内脏血管收缩,支气管和胃肠平滑肌舒张,瞳孔扩大及血糖升高等效应。这种效应常发生于劳作、危险等情况,为机体的应急反应。
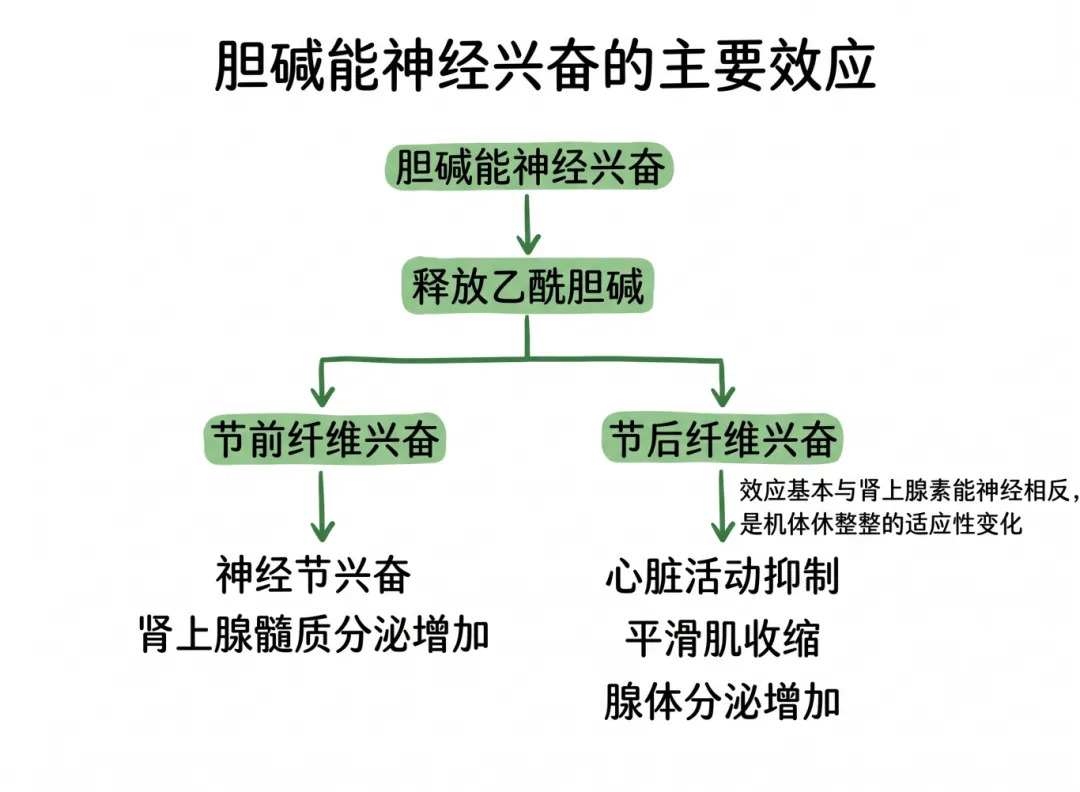
胆碱能神经兴奋时,神经末梢释放乙酰胆碱。当节前纤维兴奋时,可引起神经节兴奋和肾上腺髓质分泌增加;节后纤维兴奋时,基本上表现为与肾上腺素能神经兴奋相反的效应。这种效应常发生于静息、睡眠等情境下,是机体进行休整的适应性变化。
机体的多数器官都接受上述两类神经(胆碱能神经和肾上腺素能神经)的双重支配,而这两类神经兴奋时所产生的效应往往又相互拮抗。当两类神经同时兴奋时,则占优势张力的神经的效应通常会显现出来。如窦房结,当肾上腺素能神经兴奋时,引起心率加快;胆碱能神经兴奋时引起心率减慢,但以后者效应占优势——即两类神经同时兴奋时,则常表现为心率减慢。
上述生理作用主要为中枢神经系统功能整合的结果,主要依靠局部和整体水平的负反馈调节机制来实现。
1)局部整合:局部整合发生在ENS神经末梢突触前膜。肾上腺素能末梢释放的NA负反馈抑制NA释放的过程,即为一个典型的局部水平的调控过程,这一效应由位于突触前膜的a2受体介导。这种前膜受体结合释放的递质,继而调节该递质自身的释放过程,称为“自身受体”;递质释放的调节亦可由其他递质及其受体来介导,称为“异位受体”,在交感神经纤维末梢,M1受体、组胺受体、前列腺素受体、肽类受体等,均被发现参与NA释放的调节。
2)整体反射:整体反射包括血压调节、胃肠道运动调节、膀胱容量调节和呼吸道平滑肌的调节。其中血压调节主要依赖血管压力感受器活动引起的神经反射调节和肾素-血管紧张素-醛固酮系统的体液调节。
膨体是理解交感神经“弥散性调控”独特性的结构基础,而副交感神经末梢则采取了完全不同的布局策略。下面将从组织学结构、递质合成与储存、释放机制、受体作用、消除(降解)方式五个环节,对两者进行系统的对比阐释,并延伸到相关的病理生理过程。
(1)组织学结构:串珠状“膨体” vs 精细“终末分支”
【交感神经节后末梢(膨体型)】
交感神经节后纤维为无髓鞘C类纤维,其末梢在效应器(平滑肌、心肌、腺体)表面形成密集的神经丛,这些纤维上规律的“串珠状”膨大结构即膨体。
每个膨体并非一个经典的“终末按钮”,而是沿途释放递质的位点。一个交感神经元轴突可拥有数千至上万个膨体。膨体与靶细胞之间保持一定的间隙距离(通常20nm~数微米),膨体内富含线粒体和大量小颗粒囊泡,内含去甲肾上腺素(NE)及ATP、神经肽Y(NPY)等共递质。
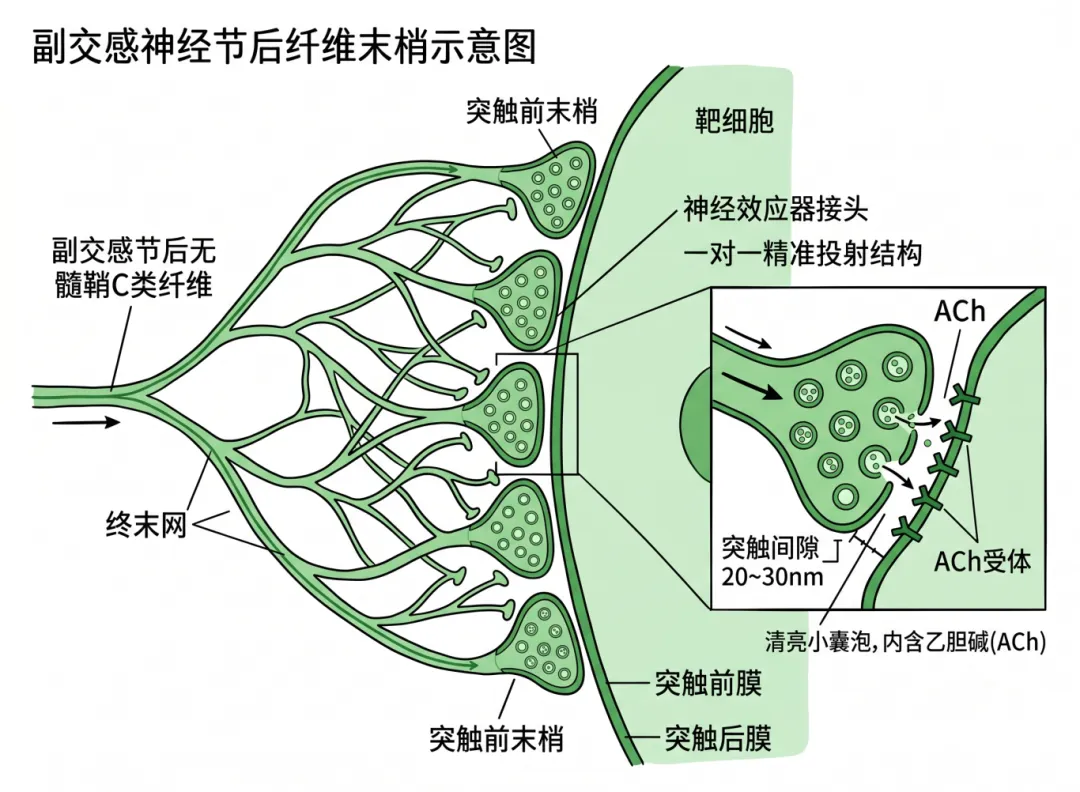
【副交感神经节后末梢(终端型)】
副交感节后纤维也是无髓鞘C类纤维,其轴突在接近效应器时,反复分支形成细密的终末网,末梢终末端直接与靶细胞膜形成典型的神经-效应器接头。终末与效应器细胞之间的突触间隙极窄(约20~30nm),类似于中枢突触的“一对一”或“一对少”精准投射。终末内主要含清亮小泡,内含乙酰胆碱(ACh)。
(2)递质的合成与储存(胞内生产线)
【交感神经(去甲肾上腺素,NE)】
合成路径(三步酶促反应):酪氨酸 →(酪氨酸羟化酶,TH,限速酶)→ 多巴 →(多巴脱羧酶)→ 多巴胺 →(多巴胺β羟化酶,DBH,存在于囊泡内)→ 去甲肾上腺素(NE)。
储存方式:NE合成后,被囊泡单胺转运体主动摄取并储存于膨体内的囊泡中,囊泡内同时高浓度储存ATP和嗜铬颗粒蛋白。
【副交感神经(乙酰胆碱,ACh)】
合成路径(一步反应):胆碱 + 乙酰辅酶A →(胆碱乙酰转移酶)→ 乙酰胆碱。
储存方式:ACh在胞质中合成后,由囊泡乙酰胆碱转运体转运并储存于突触囊泡内。其中,胆碱乙酰转移酶合成于胞体,通过轴浆运输到达末梢。
(3)递质释放机制:共同的“胞裂外排”
两者释放递质的核心分子机制高度一致,均属于钙离子依赖的胞裂外排:
去极化与钙内流:神经冲动传导至末梢时,电压门控钙通道(P/Q型或N型)开放,细胞外Ca²⁺内流,局部Ca²⁺浓度急剧升高(从10⁻⁷M升至10⁻⁴M)。
囊泡锚定与融合:Ca²⁺与突触结合蛋白结合,触发SNARE复合物构象变化,引导囊泡膜与突触前膜融合,形成融合孔。
量子释放:囊泡内容物(NE或ACh,以及共递质ATP等)通过融合孔一次性全部排入突触间隙。一个囊泡释放的递质量即为一个量子,引起微小的终板电位或接头电位。
(4)递质作用于受体(信号转导)
这是两者功能差异的核心环节,因为受体类型和后续信号通路截然不同。
(5)递质的消除(降解与回收)
【交感神经递质(NE)的消除:以“再摄取”为主】
摄取1(神经元摄取):突触前膜上的NET(去甲肾上腺素转运体)将间隙中75%~95%的NE主动重摄取回膨体末梢内。重摄取的NE可被重新包装入囊泡(储存再利用),或被胞质内的单胺氧化酶(MAO)降解。
摄取2(非神经元摄取):少量NE被效应器细胞(平滑肌、心肌)摄取,随后被细胞内的儿茶酚-O-甲基转移酶(COMT) 降解。
酶解:仅有极少部分NE在突触间隙直接被COMT降解。
病理关联1(高血压治疗):利血平通过阻断VMAT(囊泡单胺转运体),耗竭囊泡内的NE储存,导致交感传导减弱,从而降压。
病理关联2(抑郁症):三环类抗抑郁药(如丙咪嗪)通过阻断NET(去甲肾上腺素转运体),使突触间隙NE浓度升高,延长其作用时间。
【副交感神经递质(ACh)的消除:依赖“快速酶解”】
ACh的清除几乎完全依赖水解:
乙酰胆碱酯酶(AChE):位于突触间隙和突触后膜上,AChE在数毫秒内将ACh水解为胆碱和乙酸。
胆碱的再循环:水解产物胆碱被突触前膜的高亲和力胆碱转运体(CHT)重新摄取,进入末梢再次与乙酰辅酶A合成新的ACh(回收利用率极高)。
病理关联(有机磷农药中毒):有机磷化合物(如敌敌畏、沙林)与AChE的活性位点不可逆结合,使其失活。ACh无法被水解,在突触间隙持续堆积,导致副交感神经过度兴奋(瞳孔针尖样缩小、流涎、支气管痉挛、大汗、心动过缓),严重时因呼吸衰竭致死。临床急救使用解磷定重新复活AChE,并用阿托品阻断M受体以对抗堆积的ACh。
(6)递质释放的病理生理扩展:从共递质到受体失敏
【共递质的协同】
交感膨体高频放电时,囊泡除了释放NE,还释放ATP(引起早期快速收缩)和神经肽Y(NPY,强烈收缩血管并抑制NE再释放)。NPY的效应缓慢而持久,体现了“频率编码”的精细调控。
副交感末梢常与血管活性肠肽(VIP)共存(尤其在盆神经和唾液腺),VIP促进血管扩张和分泌增加。
【受体失敏与耐受】
当递质长期过度释放(如慢性心衰时NE持续升高),β1受体通过G蛋白偶联受体激酶(GRK)磷酸化并内陷,导致心脏对儿茶酚胺的敏感性下降——这是心衰时使用β受体激动剂效果差,而需用β阻滞剂“休息受体”的病理生理学依据。
【突触前自身受体负反馈】
交感末梢的α2受体可感受局部高浓度的NE,激活后抑制后续NE释放(负反馈刹车),这是机体防止过度兴奋的重要自我保护机制。
(1)紧张性支配
对效应器的支配,一般具有持久的紧张性作用,即在安静时,自主神经经常发放低频神经冲动传至效应器。
(2)功能相互拮抗
交感神经和副交感神经对同一器官的作用往往是相互拮抗。如心交感神经能加强心脏活动,而心迷走神经则起相反作用;迷走神经可促进胃肠运动和消化腺分泌,而交感神经则起抑制作用。
(3)效应器所处的功能状态对自主神经作用的影响
自主神经的外周性作用与效应器本身的功能状态有关。例如,刺激交感神经可导致动物无孕子宫的运动受到抑制,而对有孕子宫却可加强其运动。又如,胃幽门如果原来处于收缩状态,则刺激迷走神经使之舒张,如果原来处于舒张状态,则刺激迷走神经使之收缩。
(4)作用特点
交感神经的膨体串珠结构决定了其调控具有弥散性、全局性,一个冲动可以同时影响大片平滑肌细胞,适用于“战或逃”的广泛动员;而副交感末梢的紧密终末连接决定了其调控具有精准性、局部性。
节前纤维属于有髓鞘的B类神经纤维,传导速度较快;而节后纤维属于无髓鞘的C类神经纤维,传导速度较慢。故交感神经的反应弥散但启动迅速(节前纤维传导快),而副交感的局部反应则更为精准。
在递质代谢上,交感依赖“重摄取”实现节俭循环,副交感依赖“速效酶解”实现毫秒级精准制动——前者适配持久应激,后者适配精细瞬时调控。两者的结构、递质与降解方式决定了它们在昼夜节律及疾病状态下的不同表现(如有机磷中毒集中体现副交感崩溃,而嗜铬细胞瘤体现交感风暴)。
(1) 昼夜活动的规律性交替
自主神经系统的两大分支并非始终均衡活动,而是呈现出与昼夜节律相吻合的规律性交替。白天以交感神经活动占主导,机体处于“待命状态”,心率加快、血压升高、体温升高、能量释放,以适应白天活跃的工作和社交生活;夜间则以副交感神经活动占主导,心率减慢、血压和体温下降、能量储存,为睡眠和身体修复创造条件。
具体而言,早晨时段是自主神经活动切换的关键窗口。清晨时迷走神经张力降低,交感神经张力明显增高,达到一天中的活动高峰期。这种节律性变化本质上是机体为即将到来的清醒和活动所做的“预见性准备”——在真正需要用力之前,心血管系统和代谢系统已经悄悄启动了。
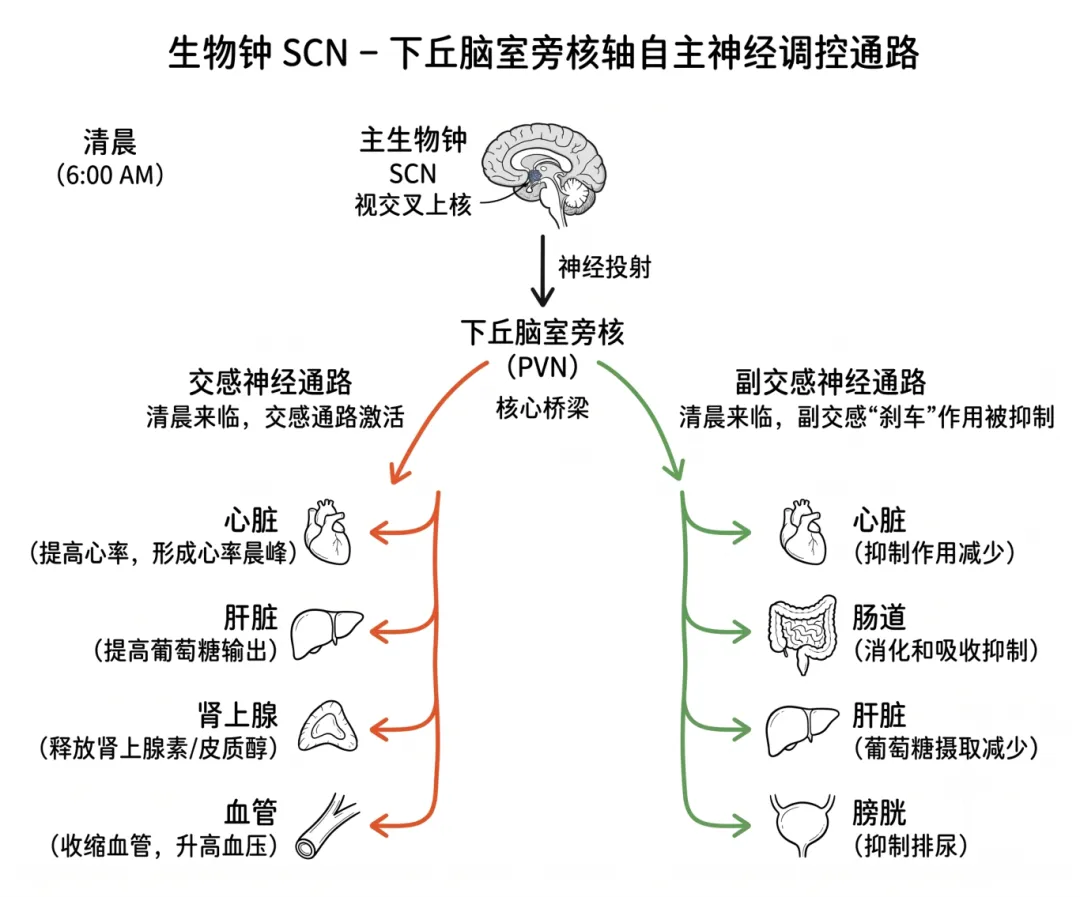
(2)生物钟(SCN)对自主神经的调控
自主神经的昼夜转化由位于下丘脑的视交叉上核(即“主生物钟”)所驱动。SCN通过两条主要通路实现对全身器官的时间调校。
第一条是神经内分泌通路:SCN通过调控下丘脑室旁核来协调激素的适时分泌,包括清晨皮质醇的上升和夜间褪黑素的释放。第二条是自主神经直接通路:SCN通过下丘脑自主神经通路直接调节器官功能,使器官能够精准地“预适应”即将到来的生理需求。
生物钟不仅在激素层面传递“时间信号”,更通过直接的神经元影响作用于全身器官,使自主神经的节律性活动成为一种全身性的、精密的协调机制。SCN-下丘脑室旁核轴在此过程中起到核心桥梁作用:一端连接着主生物钟,另一端通过自主神经系统的交感和副交感分支将“时间命令”分发到各靶器官——当清晨来临,生物钟通过交感通路提高心率和葡萄糖输出,同时抑制副交感的“刹车”作用,形成心率晨峰的自然现象。
(3) 昼夜转化带来的系列身体反应
随着交感和副交感的主导权在昼夜之间的交接,人体经历着一系列可测量的生理变化:
觉醒期:在SCN的“预适应”效应下,心率、血压、血糖和皮质醇水平的预发性上升早于活动开始;同时“黑暗激素”褪黑素下降。瞳孔扩大,呼吸道扩张以增加通气量,汗腺开始活动。
日间活动期(交感主导):心率维持较高水平,血压保持在白天应有的“工作范围”;肌肉血流量增加,基础代谢率提升;消化系统相对抑制,能量优先分配给骨骼肌和大脑;血糖水平升高以供应能量需求;情绪和精神状态处于较为警觉的水平。
夜间休息期(副交感主导):心率减慢、血压降低、体温下降;消化系统活动增强,胃肠蠕动和消化液分泌旺盛;膀胱逼尿肌的副交感促排尿作用有所增强;合成代谢占优,糖原合成和组织修复启动;瞳孔缩窄以利于休息状态的眼部舒适。
(4)节律紊乱的病理后果
当自主神经的昼夜节律被扰动时,机体多个系统将受到波及。长期熬夜、昼夜颠倒的生活方式会使交感神经在夜间应当休息时仍然高度兴奋,而副交感系统在白天该工作时却无法有效激活,造成所谓“自主神经失协调”。
在心脏层面,研究表明自主神经昼夜节律紊乱会导致心率变异性(HRV)信号的混沌特性发生变化,交感与副交感的调制强度失衡——交感调制过强时心率变异性下降,心室易颤性升高。长期睡眠剥夺还可诱发心律模式异常,增加心源性猝死风险。此外,昼夜节律的紊乱还会通过神经内分泌系统进一步放大自主神经的失调效应——在全身层面形成恶性循环。
有研究发现,SCN本身的结构性病变可能参与某些疾病的发病机制——在抑郁症或高血压患者的生物钟中已发现了病理学变化。深入来看,自主神经节律紊乱通过三条路径波及全身:直接改变各器官的神经驱动,干扰激素(皮质醇、褪黑素等)的精准定时释放,以及破坏SCN以下各级“外周生物钟”之间的时间同步,三者相互叠加,使问题从局部扩展到全身。
理解自主神经系统的昼夜节律转化,对疾病的诊断、治疗和预防具有重要临床启示。
诊断上,动态心电图记录中的HRV(心率变异性)参数能客观反映自主神经昼夜活动情况,用于评估自主神经功能状态。血压监测中血压昼夜节律的“非杓型”模式往往提示自主神经调控异常。
治疗上,降压药、抗心律失常药物和抗糖尿病药物的给药时间可依据自主神经昼夜节律进行“时辰治疗”优化。睡眠医学亦日益关注通过行为干预(规律作息、光照暴露调控、运动时间优化等)来恢复自主神经的昼夜平衡,以改善自主神经失调相关症状。心血管疾病的高危时段(清晨觉醒至上午10时)正是自主神经昼夜转化的关键窗口,也是心肌梗死、脑卒中的高发时段,这一关联更凸显了在时间生物医学框架下理解和管理自主神经系统的临床价值。
现代医学的自主神经昼夜节律,与中医的阴阳、营卫学说,是从不同视角对人体生命活动节律的深刻洞察。前者揭示了其神经生物学机制,后者则构建了一个囊括生理、病理、治疗与养生的动态平衡理论体系。
所以医生为什么总叫你要早睡呢?
如果你看到了这里,为自己点赞吧~